El termino frecuencia de gametos se aplica cuando los alelos de más de un locus se consideran. Cada gameto contiene un alelo de cada locus. Considere los dos loci A y B, los genotipos de cada locus puede ocurrir en H-W proporciones, aunque este no es el caso de los dos loci juntos. Esto puede ser de gran importancia en relación con los marcadores de genes (loci). De vez en cuando un alelo específico en el marcador loci ocurre con la enfermedad alelo de un otro locus. Si los dos loci están estrechamente ligamentados podría tener varias generaciones antes que el equilibrio sea alcanzado entre los dos loci.
La combinación de dos pares de genes A y B, cada uno con dos alelos que se muestran a continuación en el clásico dos por dos tabla, donde r, s, t, y u son las frecuencias observadas de los gametos AB, Ab, AB y ab, ver también Figura 2.5.
gen A gen B B b frecuencia
------------------------------------------------
A | r=p(A)*p(B)+D s=p(A)*q(b)-D | p(A)
a | t=q(a)*p(B)-D u=q(a)*q(b)+D | q(a)
-----------------------------------------------
Frecuencia | p(B) q(b) | 1
Un ejemplo de cálculo: El número de gametos que se observó son AB=21, Ab=49, aB=19, ab=11.
gen A/gen B B b Suma (frec)
------------------------------------------------------------
A | 21 (r=0,21) 49 (s=0,49) | 70 (p(A)=0,7)
a | 19 (t=0,19) 11 (u=0,11) | 30 (q(a)=0,3)
-------------------------------------------------------------
Suma(frec) | 40 (p(B)=0,4) 60 (q(b)=0,6) | 100 1
Las frecuencias se dan entre paréntesis: Ji-cuadrado = 9,7** Df=1
D= u-q(a)*q(b) = 0,11 - 0,3*0,6 = -0,07
Estadísticamente significativa desviación se produce a partir de la combinación
aleatoria
entre los genes A y B y corresponde a un
desequilibrio igual a -0,07.
Los gametos correspondientes procedentes de la primera tabla puede ser organizada de manera diferente como se
muestra abajo:
gametos frecuencia observada frecuencia esperada desviación
------------------------------------------------------------
AB r p(A)*p(B) D
Ab s p(A)*q(b) -D
aB t q(a)*p(B) -D
ab u q(a)*q(b) D
En la tabla se ve todos los genotipos posibles de un dos gen sistema. Recombinación física sólo han ocurrido en el doble heterocigoto, en todos los demás genotipos una recombinación no daría lugar a otros tipos de gametos diferentes de la cual se forman. El genotipo de las frecuencias correspondientes en virtud de apareamiento al azar está obtenido por la multiplicación de las frecuencias de gametos, como se muestra. La multiplicación se hace por medio de un applet diseñado para estudiar el ligamiento y el ligamiento desequilibrio.
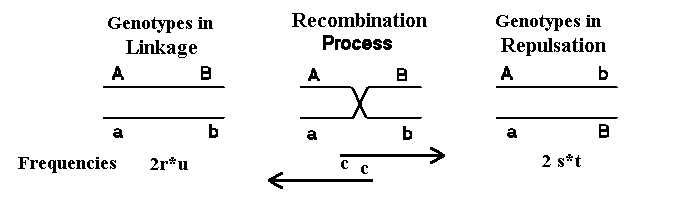
Figura 2.5 muestra los dos tipos de dobles heterocigotos.
Por la recombinación de los gametos
del otro
tipo y de forma inversa. Por lo tanto, el equilibrio de ligamiento cuando los números
son igual de los dos tipos de dobles
heterocigotos. Esto también puede ser visto en la fórmula de D dado abajo.
Figura 2.5. Los dos cromosomas homólogos en ligamiento y en
fase de repulsiones. Cuando la recombinación tiene lugar (con
una frecuencia c) se forma los gametos contrario correspondiente a la fórmula de D.
 |
La frecuencia esperada de un gameto corresponde a la frecuencia donde los dos genes son heredados independientemente. En ese caso la regla de multiplicación de probabilidades se pueden aplicar para calcular el número esperado. El desequilibrio tiene el símbolo D y se define como se muestra arriba (observados - espera frecuencia de gametos). Puede también se calcula como la mitad de la diferencia entre la doble frecuencia de heterocigotos en fase de ligamientos y repulsiones:
D = u - q(a)*q(b), o
D = r*u - t*s (= [p(AB/ab) - p(Ab/aB)]/2)
El máximo desequilibrio existe cuando todos los dobles heterocigotos están en fase de ligamiento (AB/ab) o en repulsa fase (AB/AB), entonces se tiene el valor DMax.
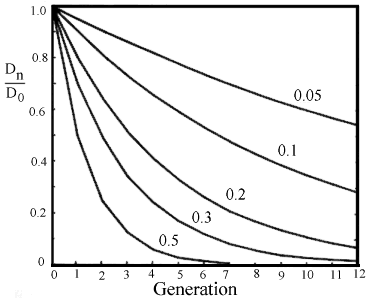
Si la frecuencia de recombinación es c, D será inflado a D*(1-c), por generación, por lo que en la generación
n Dn = D0 * (1-c) n,
donde D0 es el desequilibrio en la población base, ver figura 2.6.
Figura 2.6. Curvas de número de generaciones hacia el equilibrio de diferentes frecuencias de recombinación. |
Por dos loci independientes teniendo 0,5 en la frecuencia de recombinación de 4 a 5 generaciones pasará antes de que el equilibrio fuera alcanzado. En cuanto a loci estrechamente ligamientado con menos del 5 porciento de recombinación, que tendrá más de 25 generaciones antes de que se alcance el equilibrio.
La liberación de ligamiento desequilibrio puede producir nuevas variación genética para los rasgos cuantitativos. Si son dos loci al lado entre sí, ambos con un efecto positivo y un efecto negativo y existen en la fase de ligamiento, si surge nueva variación si los positivos alelos están recombinados y viene en fase de ligamiento
La segunda ley de Mendel predice que dos loci se segregan en forma independiente F2 en un análisis de doble cruz. Esto es causado por el hecho de que todos los individuos en F1 son dobles heterocigotos. Dado que sólo el doble heterocigotos pueden recombinarse, tomará mucho tiempo antes de equilibrio se da en una población mixta, donde la frecuencia de doble heterocigotos sólo es 2(r*u + t*s).
En relación con una nueva mutación completa desequilibrio existe en las generaciones futuras, ya que la mutación se presenta en un solo cromosoma. Este desequilibrio puede ser utilizado en relación con la aplicación de marcadores de genes para identificar la localización de los genes y de identificar los 'transportistas' en los individuos relacionados por medio del marcador loci.
Máximo desequilibrios siempre existen dentro de una familia. Esto hace que sea posible utilizar genes marcadores dentro de las familias. Si un individuo nace con una enfermedad recesiva y tiene un marcador específico combinación, los nuevos hijos que tengan la misma combinación de gen y marcador, con gran probabilidad también sean portadoras de la enfermedad.
Aplicación para el cálculo de la prueba de Ji cuadrado para el ligamiento desequilibrio, haga clic aquí, y un applet para el estudio del ligamiento y ligamiento desequilibrio haga clic aquí.