2.3 Sex-linked inheritance
Preceding page
One of the preconditions for H-W equilibrium is random mating in the population. In such a population
H-W equilibrium always occurs after one generation of random mating, provided that
both males and females have the same gene frequency.
By sex-linked inheritance there is not necessarily the same gene frequency in males and females after random mating,
since the two sexes get their genes from two different sources.
In mammals the males get all their sex-linked genes from their mothers, whereas the females get
half their genes from their sires and half from their dams. The opposite is the
case in birds, where the female is the heterogametic sex.
Example, the orange gene in cats: In a population the following number of the three
genotypes were found. The orange gene (O) gives yellow coat colour. The genotype Oo gives a mixed colour, which is caused by random
inactivation of the X-chromosome in XX individuals. The oo phenotype is non-yellow.
--------- ------ females --------- ----- males -----
Genotype OO Oo oo Total O o Total
Number 3 53 117 173 28 149 177
Frequency 0.02 0.31 0.67 = 1,00 0.16 0.84 = 1,00
----------
In mammals the sex-linked gene is shown directly in the males. That is, the genotype frequency
equals the gene
frequency. Whereas in the females the calculation of the gene frequency is identical
with the calculation for autosomal
genes, as shown below.
Frequency of O calculated as p = (2*3 + 53)/(2*173) = 0.17
do o do q = (2*117+ 53)/(2*173) = 0.83
------
1.00
There are only minor differences between the frequency in the two sexes, as
shown. If there is a difference it will
be halved in each new generation with random mating, so in practice H-W equilibrium
is reestablished after a
few generations, if the population deviated from equilibrium in the first place. Phenotypically the heterozygotes are very
special, since only one gene is active in each cell. The phenotypes occur
because of a random blend of cells with an
alternative activated gene.
From the chromosome preparation it can be seen that one of the
X-chromosome is inactive (circled), figure 2.4 left. To the
right is a heterozygotic female cat with the genotype Oo. It has yellow and black
spots caused by
alleles in the sex-linked orange locus. The white colour is caused by an autosomal gene for spots.
Figure 2.4
Heterozygote female cat with the genotype Oo in the sex-linked Orange locus. Photo Bodil Andersen, Bogense. It
gives a blended coat colour of yellow and black with variating spot sizes. This is caused by random
inactivation
of one X-chromosome in female mammals. The
inactive swine X-chromosome is circled. |
 . .
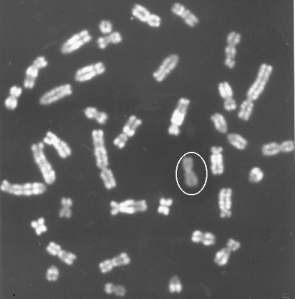 |
The inactive X-chromosome is detected by means of Acridine orange staining on
cells grown for 6.5
hours with BrdU (Brom deoxy-Uridine) in the media. DNA-syntheses which occur after adding BrdU can be seen as
weakly stained areas on the chromosomes. There is a circle around the inactive X
chromosome. The
active genes replicate early in the cell cycle and are therefore found in the white band.
The majority of cases with recessive sex-linked diseases occur in males. Haemophilia
in
human is one of the best known
examples of sex-linked recessive inheritance, the frequency in boys being 100 times
larger than the one in girls. This occurs
when the gene frequency is 0.01, corresponding to the frequency in boys. Whereas
the gene in the girls has to come
from both father and mother, each with a probability of 0.01, this corresponds to a frequency in girls of 1
in 10'000.
Next page