Figura 3.1.
Mutación,
Migración y
Selección
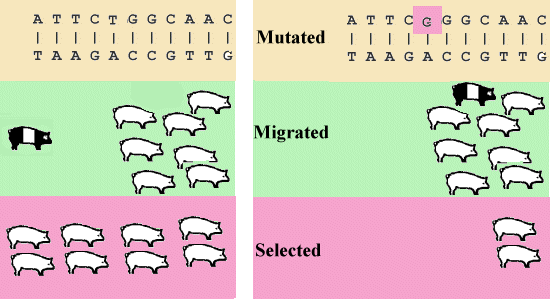
En una gran población con apareamiento al azar del equilibrio H-W ocurre a menos que la población esta sometida a efectos sistemáticos, que pueden cambiar las frecuencias génicas. Los procesos sistemáticos pueden ser divididos en las categorías mostrado en Figura 3.1.
Figura 3.1. Mutación, Migración y Selección |
|
Para los verdaderos genes (los genes de estructura) mutaciones generalmente se producen con una frecuencia de 0,1 a 1 por millón de gametos por gen. Se produce 1000 veces más a menudo en el número de repeticiones de secuencias cortas repetidas de ADN (mikrosatellites). Estas secuencias no se traducen en proteínas, pero funciona como separación entre los genes. La suma de todas las mutaciones en los genes que constituyen cada nuevo individuo lleva a una o más nuevas mutaciones. La mayoría de las nuevas mutaciones en los genes de estructura son nocivas. Por lo tanto, debido a la baja "fitness" una selección de los animales portadores de mutaciones perjudiciales se producen en cada generación. Más de un gran número de generaciones de un equilibrio entre las nuevas mutaciones y de selección se establecerá. Para genes recesivos el equilibrio frecuencia génicas (q) se puede calcular mediante la siguiente ecuación:
q2*s = my,
La frecuencia de mutación (my) es igual a la frecuencia de los individuos recesivos que son eliminados en cada generación (q2*s). Para los genes dominantes el equilibrio frecuencia génicas (p) se puede calcular mediante la siguiente ecuación:
(1/2)2pq*s = ps = my,
La frecuencia de mutación (my) es igual a la mitad de la frecuencia de animales heterocigotos, que se separa en cada generación (pq * s) y q es aproximadamente 1. Para la definición de s ver sección 3.2.
En las poblaciones naturales migración a menudo ocurre desde el barrio más cercano. Así una transición sin tropiezos en la frecuencia génicas se produce entre los sub poblaciones. En la crianza de animales migración corresponde a la introducción de nuevos animales. Normalmente se compra el mejor donde se crían los animales.
Selección natural se corresponde con el término "supervivencia del mas apto" (survival of the fittest), conocida a partir de la teoría de la evolución de Darwin. La selección natural rara vez llevan a la fijación de genes, como una población a menudo son sometidos a cambios repentinos en las condiciones ambientales. Por lo tanto, puede ser una ventaja para una población con genes que, en la situación actual, no son las óptimas. Sobre genes recesivos otra parte se puede llevar en baja frecuencia con pequeñas pérdidas de vitalidad (fitness).
El término carga de genéticos (en ingles genetic load)
es usado para describir una población con genes letales o con genes
de menores fitness. Una población sólo puede mantener una carga determinada,
que se separa con selección en cada
generación. En contraste con las nuevas mutaciones, que desempeñan un papel de menor importancia,
es más costoso mantener los sistemas, donde superdominancia ocurre con la eliminación
de parte de ambos tipos de homozigotos en cada generación.
Figura 3.2 Toro de la raza belga azul que tiene dobles músculos. El término "carga genética" se visualiza directamente. |
 |
En la raza azul belga, un mutante recesivo (músculos doble) se presenta. Esta raza se muestra en la Figura 3.2. El gen sólo se puede transmitir si un alto número de cesáreas son aceptadas como los terneros son demasiado grandes para nacer de manera natural. En la naturaleza la mayoría de estos terneros músculos dobles se mueren, por lo tanto en la naturaleza el gen sólo existe en una frecuencia muy baja.
En las secciones siguientes se dará fórmulas para predecir los cambios de frecuencias génicas cuando una población está expuesta a la presión de selección sobre un genotipo determinado.