Shows the functions of the five best known colour genes in mammals. The coat colours are caused by the melanin grains produced by the melanocytes.
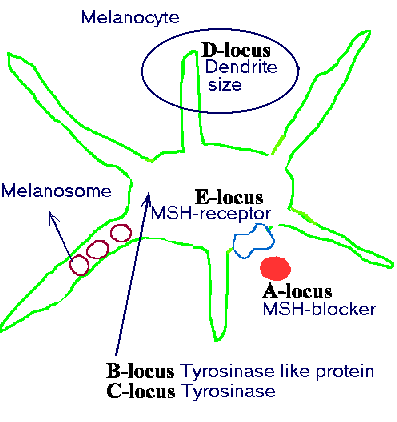
Figure 11.4 shows an illustration of a melanocyte. In the figure some of the functions of a melanocyte are pointed out. These functions are influenced by the colour genes mentioned in the last section.
MSH or the melanocyte-stimulating hormone is produced in the hypophysis. This hormone is important
in the
production of melanin. The MSH-receptor corresponds to the e-locus, whereas a blocker of this hormone corresponds
to the a-locus.
| Figure 11.4. Shows the functions of the five best known colour genes in mammals. The coat colours are caused by the melanin grains produced by the melanocytes. |
The b and c locus code for 'enzymes' which regulate the restructuring of the amino acid tyrosine, tyrosinase.
The restructuring of tyrosine is necessary for melanin to be formed.
Locus d has to do with the dendrite-size.
A comparison with the known alleles of the colour genes provides the possibility of an interpretation of their function. For instance can both a dominant and a recessive yellow exist in the agouti locus. The effect of the a and the e locus has to be strongly related, as they affect the same system, the MSH function.
The distribution of the melanocytes in the skin is also an important factor for the colour of the hair. An example of this is two gene pairs known from mice, the steel (sl or Mast cell growth factor=Mgf) and white spotting (the Kit gene), which is the Mgf receptor. These genes determine the number and the distribution of the melanocytes in the skin. They can also, as the latter gene name indicates, cause white spots. When fewer melanocytes occur in the skin the result will be a mixture of coloured and white hairs. This effect comes from the steel locus.
In mice up to 100 coat colour genes are known. They can all be categorized into one of the four classes given below:
Classifying genes which affect the coat colour:
Modification of the colour can be done by means of classical selection, where the intensity of the colour is treated as a quantitative character. For instance, the colour of wild mink is dark brown. By selection for darker colour through 40 generations a completely black mink colour type has been created.