The Landrace and Yorkshire are homozygote for dominant white, the Duroc is red and the Hampshire is black with a white belt

The inheritance of coat colour in the Danish swine breeds are regulated by the following three loci, see also Figure 11.5.
Dominant Recessive ------------------------------------------------- White I- Coloured ii Black E- Red ee Uniformly ii White belt i(be)i(be) -------------------------------------------------
The Landrace (L) and the Yorkshire (Y) breeds are homozygotic for dominant
white, II.
The Duroc (D) race is red, eeII.
The Hampshire (H) race is black with a white belt, EEi(be)i(be).
| Figure 11.5 The Landrace and Yorkshire are homozygote for dominant white, the Duroc is red and the Hampshire is black with a white belt |
|
Most of the pigs produced in Denmark have an LY mother and a coloured (D or HD) father. They are all white as they are heterozygote with respect to the genotype Ii. Marklund et al. 1998, Genome Res. 8:826-33, has shown that the dominant white in swine is caused by an allele in the kit gene, which corresponds to the mast cell's growth factor receptor. The extension locus code for the colour difference between the black and red pigs.
Details of the belt gene is given in Mamm Genome 1999 Dec;10:1132-6 af Giuffra E et al. it is an allele in the kit gene.The inheritance of coat colour in the Danish cattle breeds are regulated by the following three loci: `
Dominant Recessive --------------------------------------- Black E- Red ee Uniformly S- Spotted ss White head SH- Uniformly SS ----------------------------------------
The Danish Holstein Frisian dairy breed (SDM) has the genotype EEss and The Danish
Red dairy breed (RDM) has
the genotype eeSS. Crosses between SDM and RDM become uniformly black. Some RDM have
a different colour pattern,
tiger stripes, which are cased by an allele in the e locus which dominates over red colour.
In some Danish beef cattle breeds, for instance Hereford, white head colour pattern
occur, having dominant
inheritance. It is inherited
from a dominant allele in the same locus as spotted. It is known, that the gene for
spotted
is situated on chromosome 6, and is tightly linked with the kit gene
and it has interaction with the kit gene.
| Figure 11.6. Danish Holstein Frisian with the genotype EEss and Danish Red with the genotype eeSS. The Hereford has the dominantly inherited white head.  |
Klungland et al. 1995, Mammalian Genome 6:636-39, have shown that red colour in cattle is caused by the ee genotype in the extension locus.
Inheritance of coat colour in dogs,
by Helle Friis Proschowsky
The inheritance of coat colour in dogs is very complex, therefore only the main types will be described
here.
In some breeds the coat colour is very constant, thus all individuals have the same coat colour. But each breed has its
own special set of colour genes and some breeds have great variation. People with special interested in a particular
breed can consult 'The inheritance of coat colours in dogs' by Clarence C. Little, Howell Book House, 1984.
A short description of the best known colour loci and their significance for the coat colour in dogs:
Locus A: Determines the amount and the localization of dark and light pigment, both for the individual
strands of hair and for
the fur as a whole. The allele at , for instance, causes the black and tan colour
of Rottweilers, Airedales and Gordon setters.
Locus B: Determines the amount of dark pigment and whether the colour should be dark (B-) or liver brown (bb).
Locus C: Determines the depth of the pigmentation. The allele ca
, gives albinos, and the chinchilla allele cch which makes the
reddish and yellow colours become cream colour in for instance Cocker Spaniel. The chinchilla allele also gives rise to
large variations in the depth of the pigmentation, in for instance Chesapeake bay retriever and Tibetan Terrier.
Locus D: Determines the
intensity of the colour. The allele D gives an intense colour (black in Grand
Danois), while the allele d gives a bluish
dilution. Weimarans always carry the Recessive dd along with bb, thereby yielding the liver brown coat colour.
Locus E:
Determines the pattern of the pigmentation in the coat. Alleles Em, for instance gives
the Leonberger a black mask and the
allele eb gives the Boxer and the Grand Danoi tiger strips (brindle), while the genotype ee
produces the Labrador Retriever's yellow colour.
Figure 11.7 The black and tan colour in the Airedale, atat.
Dilution colour in the Tibetan, dd, and yellow colour from the
extension locus in the Labrador retriever,
ee.

|
Locus G: alleles in this locus determine whether or not the colour fades with age.
Locus M: the dominant allele M gives
merle colour (Collie, Shetland Sheepdog), while the Recessive allele m gives a
uniform pigmentation. It is well known
that two dogs carrying the merle colour should not be mated, as a double dose of
the allele often causes damage to the eyes
and the sense of hearing.
Locus S: determines whether or not the coat
have spots. The dominant allele S gives a uniform pigmentation,
though occasionally with white spots on the feet and/or the chest (Retriever, Boxer and New
Foundlander). The three
more Recessive s-alleles give varying amounts of white spots, the Clumber Spaniel and
the Dalmatian being mostly white.
Inheritance of coat colour in the cat, by Nan Hampton, University of Texas at Austin, who also has an elaborated power point show with about 50 slides.
Below is shown a table of some of the more common genotypes and phenotypes of the domestic cat.
| Common Genotypes and Phenotypes | |||||
| Homozygous | Homozygous | Heterozygous | |||
| Genotype | Phenotype | Genotype | Phenotype | Genotype | Phenotype |
| AA | agouti | aa | non-agouti (black) | Aa | agouti |
| BB | black pigment | bb | brown pigment | Bb | black pigment |
| CC | full color | cbcbcscs | burmesesiamese | CcbCcscbcs | full colorfull colortonkanese |
| DD | dense pigment | dd | diluted pigment | Dd | dense pigment |
| ii | full pigment | II | pigment limited to tips of hairstrand, base of hair light gray or white, agouti especially affected by agouti=silver tabby with non-agouti=smoke with orange=cameo | ||
| LL | short hair | ll | long hair | Ll | short hair |
| mm | long tail | MM | lethal on prenetal stage | Mm | short tail |
| oo or oY | non-orange (usually black) | OO or OY | orange | Oo | tortoiseshell=orange and black in females |
| ss | no white spots | SS | white spots > 1/2 body | Ss | white spots < 1/2 body |
| TT | mackerel stripe | TaTatbtb | Abyssinianblotched | TTaTtbTatb | Abyssinian, faint leg and tail stripes Mackerel stripes Abyssinian, faint leg and tail stripes |
| ww | not all white | WW | all white | Ww | all white |
Inheritance of coat colour in mink
Mink's colour types have evolved through selection or mutations of colour genes. The wild genotype for colour, which
has no mutations, is called Standard and ranges from dark brown (wild mink found in
nature and the
colour type Wild mink) to the strongly selected farm mink Black standard, which is totally black. This is due to
selection for darker colour in the last 40 generations.
The mutated colour types in mink are classified according to numbers and types of genes responsible for the appearance of the colour. Until now it has not been possible to correlate the mink colour genes with the colour genes described in other mammals. The only exception is the albino locus. Hopefully this problem will be solved in the near future by means of DNA technology.
The Recessive and dominant colour genes: Single Recessive colours stem from homozygosity in mutated Recessive genes. Colour genes from two or more mutated colours, having originated from the same wild (standard) genes, are called allelic genes and will occupy the same chromosomal location (locus). The homozygous state of dominating genes is often lethal, resulting in a reduced litter size when heterozygotes are mated. Only the most commonly used combinations of the mutations are shown. A cross between non-allelic mutations results in the dark brown colour. This was the common type when the mutations originally were established.
| Colour types, single | Mutation name | Mutation genotype | Wild type, Standard |
| White | Albino, red eyeds | cc | CC |
| Hedlund White, deaf | hh | HH | |
| Grey | Aleutian, bluish | aa | AA |
| Silver Blue, grey | pp | PP | |
| Steel Blue | psps | PP | |
| Brown | Royal Pastel | bb | BB |
| Moyl | mm | MM | |
| American Palomino | kk | KK | |
| Bicoloured | Black Cross | Ss | ss |
| 95 % White | SS | ss | |
| Finn Jaguar | Zz | zz | |
| Combinations | |||
| Bicoloured | Pastel Cross | bb Ss | BBss |
| Bluish grey | Sapphire | aa pp | AAPP |
| Blue Iris | aa psps | AAPP | |
| Sand | Pearl | mmpp | MMPP |
| Light sand | Violet | aakkpp | AAKKPP |
Combination types used as production crosses. The crossing of non-allelic Recessive genotypes is practiced to produce various dark brown shades (called Demi buff, Glow or Wild mink).
The following three crosses (F1) are commonly used to produce the Glow type
Sapphire x Pastel
Pearl x "
Violet x "
F1 x Wild mink
The Pastel is the female due to its high fertility. The F1 female can be crossed with Wild mink. The F1 female is very fertile and the offspring will maintain the Glow colour type as the Wild mink breed true for the Standard genes. The Mahogany colour type is produced by crossing Standard black and Wild mink. This gives a very dark brown mahogany colour type. The Mahogany F1 can be used in further breeding with the same result, therefore a pure bred Mahogany line can be established.
References: Experience from farm practise.
Nes, N., et al. 1988. Beautiful Fur Animals and their Colour Genetics. Scientifur, 60 Langagervej, DK-2600 Glostrup.

Colour genes in the horse
For a detailed description of the horse colour genes reference can be made to
'The horse coat colour genetics' from the Model Horse Reference website.
Horse Colours Genetics
or from the Veterinary
faculty in Davis, California
Horse Colours
The most important horse colour genes with indication of back ground colour genes :
| Agouti locus (A locus) | back ground CCEE | ||||
| Dominant bay, A- |  |
Recessive black aa |  |
||
| Cremello locus (Cr locus) | back ground aaee | ||||
| 'Codominant' chestnut, CC |  |
Heterozygot, CCr | 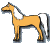 |
'Recessive' light, CrCr | 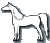 |
| Extension locus (E locus) | back ground aaCC | ||||
| Dominant black, E- | |
Recessive chestnut, ee | |
||
| Grey locus (G locus) | back ground AACCEE | ||||
| Dominant grey, G- |  |
Recessive bay, gg | |
||
| Roan locus (R locus) | back ground AACCEE | ||||
| Heterozygot roan Rr |  |
Recessive bay, rr | |
Dominant RR, die | |
| Dominant white (W locus) | back ground AACCEE | ||||
| Heterozygot white Ww | |
Recessiv bay, ww | |
Dominant WW, die |
The colour genes work in concert, the types shown above are
representative for a group, in the real world there is a continuum of
colours due to modifying genes.
The genes in the agouti and extension locus works as earlier mentioned together,
so if a horse should be black it shall have the genotype aaE-; an aaee horse
has chestnut colour.
There is also interaction between all other colour genes.
Roan and ekstension locus are linked.
Locke et al. (2001)
Animal Genetics 32:340-343, have shown
that the cream dilution, Cr, allele is not coded from the C locus,
as earlier symbols have indicated. The gene is not homologous
with the tyrosinase gene in other species.
Other species
The rabbit's coat colour genetics reference is to Jackie Carey's
Rabbit Colour Genetics