Figure 3.1.
Mutation,
Migration and
Selection
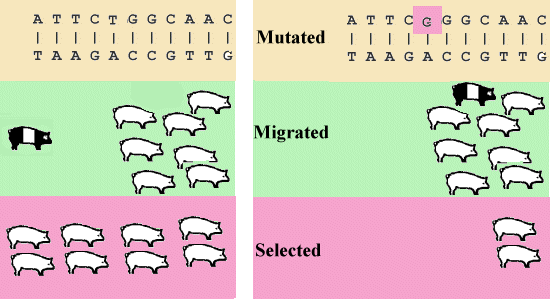
In a large population with random mating H-W equilibrium occur unless the population is subjected to systematic effects, which can change the gene frequency. The systematic processes can be divided into the categories show in Figure 3.1.
Figure 3.1. Mutation, Migration and Selection |
|
For the proper genes (structure genes) mutations generally occur with a frequency of .1 to 1 per million gametes per gene. It occurs 1000 times more often in the number of repetitions in short repeated DNA sequences (mikrosatellites). These sequences are not translated into protein but works as internal spacers between the genes. The sum of mutations in all genes will cause each new individual to carry one or more new mutations. Most new mutations in the structure genes are harmful. Therefore due to low fitness a selection of animals carrying the harmful mutations will occur in every generation. Over a large number of generations a balance between new mutations and selection will be established. For recessive genes the equilibrium gene frequency (q) can be calculated by the following equation:
q2*s = my,
the mutation frequency (my) is equal to the frequency of recessive individuals being removed in each generation (q2*s). For dominant genes the equilibrium gene frequency (p) can be calculated from the following equation:
(1/2)2pq*s = ps = my,
the mutation frequency (my) is equal to half of the frequency for heterozygotic individuals, which is removed in each generation (pq*s) and q is approximately 1. For definition of s see section 3.2.
In natural populations migration often occurs from the closest neighbourhood. Thereby a smooth transition in the gene frequency will occur between the sub populations. In animal breeding migration corresponds to introduction of new animals. They are normally bought where the best animals are bred.
Natural selection corresponds to the term 'survival of the fittest', known from Darwin's theory of evolution. The natural selection will seldom lead to gene fixation, as a population often are subjected to sudden changes in environmental conditions. Therefore it can be an advantage for a population to carry genes which, in the present situation, are not the optimal. Recessive genes on the other hand can be carried in low frequency with only minor loss of vitality (fitness).
The term genetic load is used to describe a population with
'lethal genes' or or genes with lower fitness.A population can only carry a
certain load which is removed by selection in each generation. In contrast to
new mutations, which play a minor role, it is more
costly to maintain systems, where overdominance occurs by removal
or partly removal of both homozygotic types in each generation.
Figure 3.2 Bull of the Belgian Blue breed which is double muscled. The term 'genetic load' is visualized. |
 |
In the Belgian Blue breed a recessive mutant (double muscles) is found. This breed is shown in Figure 3.2. The gene can only be passed on if a high number of Cesarian sections are accepted as the calves are too big to be born the natural way. In nature most of these calves would die, therefore in nature the gene would only exist in a very low frequency.
In the next sections formulas will be given for how the gene frequencies change when a population is exposed to selection pressure on a specific genotype.