Der findes i litteraturen talrige beretninger om vellykket selektion mod arveligt betingede sygdomme; men i næsten alle tilfælde drejer det sig om simpelt recessive egenskaber, som bekæmpes ret rutinemæssigt. Dog er der naturligvis problemer i de tilfælde, hvor den iværksatte selektion modvirkes således som man kommer ud for i forbindelse med overdominansfænomenet, hvor heterozygoterne har en større reproduktionsegnethed (fitness) end homozygoterne.
Når der derimod er tale om sygdomme med en polygenisk disposition, er beretninger om heldigt udført selektion sparsomme for de større husdyrs vedkommende. De resultater, der er opnået i retning af større sygdomsresistens, er for en stor del baseret på krydsning. At det ikke blot i teorien, men også i praksis er muligt at opnå væsentlige resultater af selektion mod specifikke sygdomme, er med stor tydelighed vist indenfor hønseavlen og ved avl med laboratoriedyr.
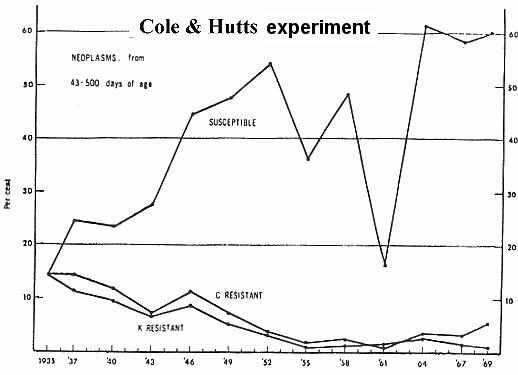
| Figur 12.3. Selektions forsøg for og imod neoplasma dødsfald hos høns. |
|
Initialgenerationen var uselekteret, men for hver af de følgende generationer udvalgtes resistente avlsdyr i hver linje. I 1935 etableredes tillige en tredje linje, S, på grundlag af individer, der var modtagelige (susceptible line) for leukose. Denne linje blev i alle følgende generationer vedligeholdt ved fortsat selektion af avlsdyr, der var modtagelige. De tre linjer opholdt sig i fælles miljø. Bortset fra perioden 1958-61 (jf. grafen) må miljøet antages at have været virusinficeret i betydelig grad i hele forsøgsperioden. Figuren viser tydeligt, at selektionen har været ret effektiv i begge retninger, idet der ved forsøgets afslutning (opgørelse 1967-69) var en neoplasme -dødelighed på ca. 60 % i den modtagelige linje, medens de to resistente linjer, C og K, havde en neoplasme-dødelighed på henholdsvis 3,7 % og 0,9 %. I samme periode reduceredes det totale antal dødsfald fra 51,5 % (C) og 44,4 % (K) til henholdsvis 12,7 % og 8,0 %. Dødsfaldene er opgjort i perioden fra 43 til 500 dage efter klækningen. De nævnte selektionsresultater er opnået til trods for forholdsvise lave heritabilitetsestimater og til trods for, at der samtidig blev selekteret for mange andre egenskaber. De gode resultater må dog forklares med, at der er anvendt afkomsundersøgelser som udvalgskriterium. I øvrigt mener Cole & Hutt, at samme resultater kunne være opnået på betydeligt kortere tid under anvendelse af en selektionsprocedure, som kun blev anvendt i en del af forsøgsperioden.
Da leukocyter er af betydning ved udvikling af resistens, er det af interesse at vide, at det er muligt at selektere for højt og lavt leukocyttal i blodet (WBC= white blood cell). Sådanne forsøg er udført hos mus (Chai 1975, J.Heredity 66:301-309). I initial populationen, (generation 0) havde man et leukocyttal på 6-8 x 103 pr. mm3. Efter 22 generationers selektion i HLC (high leucocyte count) og LLC (low leucocyte count) linjerne nået tallene henholdsvis 36-38 x 103 og 4-5 x 103, medens kontrolpopulationen har nået et niveau på ca . 10 x 103 , altså ikke væsentligt forskelligt fra initialpopulationens (en evt. signifikant forskel kan bero på "random genetic drift").
I relation til dette selektionsforsøg skal til sidst nævnes, at man har beregnet en h2 værdi på 0,20 for lymfocyttallet hos visse kvægracer. Hvis det er ønskeligt, kan lymfocyttallet altså let ændres ved selektion.
| Figur 12.4. Selektions forsøg for højt og for lavt antistof titer mod fåre erotrocytter hos mus. |
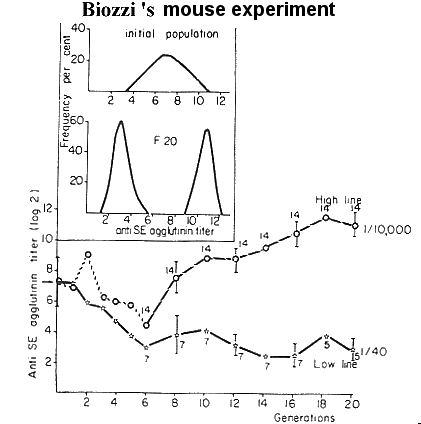 |
For at afprøve de af Biozzi et al. (1972, J.Exp.Med. 135:1071-1094) selekteret mus for højt og lavt antistofrespons i forbindelse med experimentel immunisering, er der gennemført en lang række challenge forsøg med experimentelle infektioner på de to muselinier. Det skal her bemærkes at de mus der har et højt generelt antistofsvar generelt har et dårligt cellulært forsvar. Individer med godt cellulært forsvar vil fjerne antigenerne v.h.a. deres makrofager, og derfor vil antigenet blive fjernet, før der kan dannes antistof mod det. Så det naturen har fundet frem til er et kompromis mellem et godt cellulært og humoralt immunforsvar, som det også fremgår af figur 12.5.
| Figur 12.5. Challenge forsøg med forskellige mikroorganismer på dyr selekteret for højt og for lavt antistof titter mod fåre erotrocytter hos mus. |
 |
Konklusion:
Den konklusion som kan drages fra de gamle forsøg må være, at det er muligt at avle for speciel
resistens, men en sådan avl vil normalt blive overhalet af muligheden for vaccination, hvilket
også er sket for hønseleukose vedkommende. Det må endvidere konkluderes, at det ikke er
muligt at selektere for generel resistens ved at selektere for høje eller lave værdier af
immunitetens komponenter, idet de er modsat rettede og deres naturlige niveau vil være
balanceret i forhold til en populations historie med hensyn til infektiøse agens. Så renavl kan
ikke benyttes til at forbedre den generelle resistens
baseret på de immunologiske komponenter. Men hvad med krydsning?
Ved at se på Biozzi's graf fra callenge forsøget kan man konkludere,
at det alt andet lige er bedst at ligge i
nærheden af middelværdien, idet det er de ekstreme dyr,
der udsættes for de sværeste sygdomsangreb. Så et af midlerne til at opnå så
lav sygelighed som muligt er at
arbejde med en ensartet population af dyr med den optimale middelværdi.
Ensartethed kan opnås via krydsningsdyr. Tovejs, trevejs eller
tilbagekrydsninger er på grund af dominansforholdene mere ensartede end renavlsdyr.
Det skulle derfor være muligt at finde frem til krydsningskombinationer, der
har den rigtige middelværdi i
forhold til en populationens sygdomsudfordringer.
F2 dyr eller fire-vejskrydsninger er mere
heterogene, og ydergrupperne vil derfor, som i renavlspopulationer,
være mere udsatte for forskellige sygdomme, så
anvendelse af den type dyr i produktionen bør normalt undgås.
Der foreligger p.t. ingen praktiske forsøg, der kan verificere de fremsatte postulater
vedr. forskel i sygelighed mellem de forskellige krydsningstyper. Der bør derfor snarest
iværksættes analyser af disse forhold.