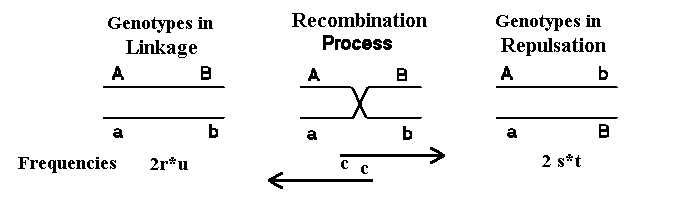
Figur 2.5. Viser de to homologe kromosomer hos de to mulige heterozygoter i de to modsatte faser. Sker der rekombination dannes den modsattes gameter. Dette sker med frekvensen c
Gamet- eller haplotype-frekvenser anvendes, når man ser på to loci samtidig. Hver ny gamet vil indeholde en allel fra hvert locus. Ser man på to loci A og B, kan der inden for hvert loci være H-W-ligevægt, uden at det er tilfældet for de to loci tilsammen. Dette kan have stor betydning i forbindelse med DNA-markører, hvor det kan ske, at en bestemt allel altid forefindes sammen med en skadelig allel på det andet loci. Er de to loci tæt koblede, vil det vare mange generationer, før der opstår ligevægt mellem de to loci.
Definition af koblingsuligevægt og statistisk test: Vi vil nu se på de mulige gameter, der kan dannes ud fra to loci A og B med hver to alleler. Kombination af to loci A og B med hver to alleler er vist nedenfor i en klassisk tabel, hvor r, s, t, og u er de observerede gametfrekvenser af gameterne AB, Ab, aB og ab. Gameterne er opstillet i tabel som kendt fra den almene genetik, de forventede frekvenser svarer til sandsynlighedsregningens multiplikationsregel:
gamet observeret frekvens forventet frekvens afvigelse
-----------------------------------------------------------------
AB r p(A)*p(B) D
Ab s p(A)*q(b) -D
aB t q(a)*p(B) -D
ab u q(a)*q(b) D
Eksempel med statistisk test: Det observerede antal gameter er følgende: AB 6, Ab 49, aB 34 og ab 11. Den statistiske test for koblingsuligevægt illustreres bedst når tallene indsættes i den klassiske to gange to tabel med test for uafhængighed svarende til D=0. Gamet og allel-frekvenserne er anført i parenteser.
locus A/locus B B b Sum (Frek)
------------------------------------------------------------
A | 6 (r=0,06) 49 (s=0,49) | 55 (p(A)=0,55)
a | 34 (t=0,34) 11 (u=0,11) | 45 q(a)=0,45)
-------------------------------------------------------------
Sum (Frek) | 40 (p(B)=0,4) 60 (q(b)=0.6) | 100 1
Chi-i-anden = 43.1*** Df=1
D= u-q(a)*q(b)=0,11 - 0,45*0,6= -0,16
Der er statistisk signifikant afvigelse fra tilfældig kombination mellem generne A og B svarende til en disekvilibrium på -0.16. I tilfælde af en sygdomsallel store B er chancerne for at have den meget stor (34/40) når der er et lille a allel i lokus A.
Detekterbare rekombinanter: I nedenstående skema er vist de mulige genotyper i et to loci system. Dobbelt heterozygoterne er fremhævet med fed skrift. Fysisk rekombination har kun betydning hos dobbelt heterozygoterne, hos alle andre genotyper vil fysisk rekombination ikke give anledning til detekterbar rekombination. De tilsvarende forventede genotypefrekvenser under tilfældig parring fås som vist ved multiplikation af gametfrekvenserne.
gamet mor AB Ab aB ab r s t u far ------------------------------------ AB | AABB AABb AaBB AaBb r | r*r r*s r*t r*u Ab | AABb AAbb AaBb Aabb s | s*r s*s s*t s*u aB | AaBB AaBb aaBB aaBb t | t*r t*s t*t t*u ab | AaBb Aabb aaBb aabb u | u*r u*s u*t u*u
I figur 2.5 er vist de to typer dobbelt heterozygoter AB/ab og Ab/aB. Ved rekombination dannes gameter af den andens type og omvendt. Der er derfor koblingsligevægt, når der er lige mange af hver af de to typer dobbelt heterozygoter.
Dette udmønter sig i en formel mere for D som vist nedenstående.
Figur 2.5. Viser de to homologe kromosomer hos de to mulige heterozygoter i de to modsatte faser. Sker der rekombination dannes den modsattes gameter. Dette sker med frekvensen c |
|
D = u - q(a)*q(b), eller
D = r*u - t*s (= [f(AB/ab) - f(Ab/aB)]/2)
Den maksimale uligevægt opstår, når alle dobbelt heterozygoter er i samme fase (AB/ab) eller (Ab/aB); den har da værdien DMax.
Er rekombinationsfrekvensen c, vil D formindskes til D*(1-c) per generation, så man har
i n'te generation
Dn = D0*(1-c)n, hvor
D0 er uligevægten i udgangspopulationen, se figur 2.6.
Figur 2.6. Kurver over antal generationer hen mod koblings-ligevægt for forskellige rekombinations frekvenser. |
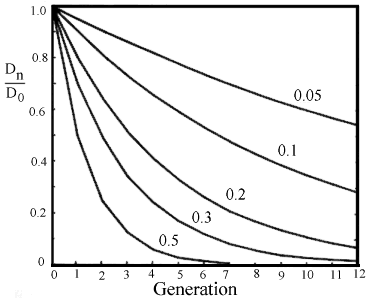 |
For to uafhængige loci med en rekombinationsfrekvens på 0,5 går der 4 til 5 generationer, før der i praksis opstår ligevægt. Drejer det sig derimod om tæt koblede loci med under 5 % rekombination, går der let 25 generationer, før man nærmer sig ligevægt, se Figur 2.6.
Der er som vist ovenstående i alt 16 genotype-kombinationer, når man trækker en tilfældig gamet fra henholdsvis far og mor hvilket svarer til de klassiske Mendelske to genspaltninger. Mendels anden lov udsiger, at der opstår koblingsligevægt i F2 i en analysekrydsning, hvilket skyldes at alle individer i F1 er dobbelt heterozygoter i den ene fase. Da det kun er dobbelt-heterozygoterne, der kan 'rekombinere', går det meget langsommere i en blandingspopulation, hvor frekvensen af dobbelt-heterozygoten kun er 2(r*u + t*s).
Opbrud af koblingsuligevægt kan give anledning til ny genetisk variation for kvantitative egenskaber. Er to gener koblede, og har de henholdsvis positiv og negativ effekt, og de sidder på samme kromosom, opstår den nye variation ved, at rekombinerer bringer de to positive gener til at sidde på samme kromosom.
I forbindelse med en ny mutation er der koblingsuligevægt i mange generationer fremover, da mutationen opstår på et og kun et kromosom. Denne koblingsuligevægt kan bruges i forbindelse med anvendelse af DNA-markører til påvisning af genets placering på et unikt kromosom og til påvisning af anlægsbærer ved hjælp af DNA-markører.
Der er altid maksimal koblingsuligevægt indenfor en familie. Dette gør det muligt at arbejde med DNA-markører indenfor familier. Er der født et sygt individ med en recessiv sygdom types dette med DNA-markørerne. Er der tæt kobling vil fremtidige afkom med samme kombination af DNA-markører med stor sandsynlighed også være syge.
Applet til beregning af Chi-i-anden for koblingsligevægt, klik her, og Applet til studie af kobling og koblingsuligevægt, klik her.