Mutation,
Migration og
Selektion
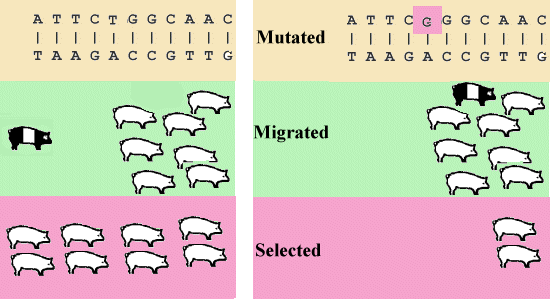
Både mutationer og migration introducerer nye alleler i en population.
Mutationsraterne er almindeligvis lave, derfor bidrager mutation kun lidt
til ændring af allel frekvenser. Naturlig og menneskeskabt selektion kan
ændre allel frekvenser på en systematisk måde til fordel for en af allelerne
(genotyperne). Selektion kan også resultere i en stabil genetisk polymorfi, hvor
heterozygoten har den højeste fitness. Selektion mod skadelige mutationer kan
føre frem til en selektions-mutations balance, hvor frekvensen af den skadelige
allel forbliver konstant fra generation til generation. Den lille population er
ustabil pga. 'genetisk drift', der er forårsaget af tilfældige ændringer af
genfrekvensen. Retningen på ændringen er tilfældig, og kan være meget hurtig
i små populationer på under 100 avlsindivider.
-----
I en stor population med tilfældig parring er der H-W ligevægt, med mindre populationen er underkastet systematiske påvirkninger, der kan ændre genfrekvensen. De systematiske processer kan opdeles i følgende kategorier.
|
Figur 3.1.
Mutation, Migration og Selektion |
|
Mutation sker normalt med en frekvens på 1 per million gameter per gen for de egentlige arveanlæg (strukturgener). Den sker ca. 1000 gange hyppigere, når det drejer sig om antal repetitioner i korte repeterede DNA sekvenser (mikrosatelliter). Disse sekvenser bliver ikke oversat til et protein, men virker som interne spacere mellem generne. Summen af mutationer over alle arveanlæg vil bevirke, at hvert nyt individ bærer på en eller flere nymutationer. De fleste nymutationer i strukturgenerne er skadelige, derfor sker der i hver generation en vis selektion på grund af lav fitness hos dyr, der er bære af de nye skadelige mutationer. Der vil over et stort antal generationer opstå en balance mellem nymutation og selektion.
For recessive arveanlæg kan ligevægtsgenfrekvensen (q) beregnes fra følgende ligning
q2*s = m,
hvor mutationsfrekvensen (m) er lig frekvensen af recessive individer, der bortselekteres i hver generation (q2*s).
For dominante arveanlæg kan ligevægtsgenfrekvensen (p) beregnes fra følgende ligning
(1/2)2pq*s = ps = m,
hvor mutationsfrekvensen (m) er lig med halvdelen af frekvensen af heterozygote individer, der bortselekteres i hver generation (pq*s) og q er approximativ 1. For definition af s se afsnit 3.2.
Migration sker i naturlige populationer oftest fra de nærmeste omgivelser. Derved får man jævne overgange i genfrekvens mellem de enkelte subpopulationer. I husdyravlen svarer migration til indkøb af avlsdyr, disse bliver normalt købt hvor de bedste dyr forefindes.
Naturlig selektion svarer til begrebet 'survival of the fittest' der er kendt fra Darwins udviklingslære. Den naturlige selektion vil sjældent nå til genfiksering, da en population i visse tilfælde pludseligt får ændrede milieuforhold. Derfor kan det være en fordel for populationen at bære på gener der ikke i den øjeblikkelige situation er de mest optimale. Når der er tale om recessive gener, kan disse bæres i lav frekvens med ringe tab af livskraft (fitness) til følge.
I forbindelse med nedsat fitness eller letalgener taler man om 'genetic
load' for populationen. Populationen taber de bortselekterede i hver generation
og en population er kun i stand til at bære en vis byrde, her spiller
nymutationerne dog kun en mindre rolle. Det er derimod en meget stor omkostning
at vedligeholde systemer, hvor der er overdominans med bortselektion af
begge de homozygote typer med en vis frekvens i hver generation.
Figur 3.2. Tyr af Belgisk Blåhvid race med dobbelt muskelhed. |
 |
I de næste afsnit vil der blive givet formler for hvorledes genfrekvensen
ændre sig, når der er lagt et selektionstryk på en bestemt
genotype. Formlerne er udviklet, så de både kan anvendes ved
naturlig såvel som menneskeskabt selektion på begge køn.
I husdyrpopulationer behandles de to køn vidt forskelligt, derfor
skal formlerne ofte anvendes på hvert køn for sig. Har man
f.eks. en situation hvor man kan DNA teste handyrene for en recessiv
nedarvet sygdom, og dermed sikre at alle avlshannerne er fri for sygdomsgenet.
Da vil man halvere frekvensen af sygdomsgenet i hver generation, da halvdelen
af generne kommer fra fædrene og halvdelen fra mødrene.