Ved selektion mod den recessive får man følgende opstilling. Selektion (s) mod den recessive skal opfattes relativt i forhold til de dominante typer. Lille s er den proportion af en given genotype, der bortselekteres i hver generation.
Ved selektion får man: --------- Genotype EE Ee ee Total Frekvens p2 2pq q2 = 1,00 Fitness 1 1 1-s Proportion p2 2pq q2(1-s) = 1-sq2 efter selektion ---------
Efter selektion beregnes genfrekvensen ved gentællemetoden, q' bestemmes som halvdelen af heterozygoterne plus de overlevende recessive relativ til halvdelen af proportionen efter selektion af alle overlevende gener, der er lig 1-sq2.
q' = (2pq/2 + q2(1-s))/(1-sq2)
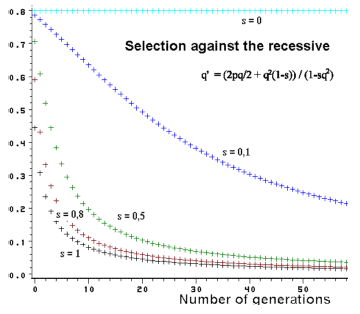
| Figur 3.3. Selektion mod den recessive og ændring i genfrekvens over generationerne. Ved lave genfrekvenser er ændringerne meget langsomme. |
|
I figur 3.3 kan man se en sådan anvendelse. Ved stærk selektion, s=1, ændres genfrekvensen meget hurtigt ved høje genfrekvenser. Er genfrekvensen derimod lav, påvirkes frekvensen kun i ringe grad. Populationen parkerer genet i en slags venteposition. Dette er meget ubehageligt, såfremt det drejer sig om et sygdomsgen i en husdyrbestand. Man bliver aldrig helt af med det skadelige gen. Der er derfor stor interesse for at finde DNA testmetoder, hvorved man kan påvise heterozygote anlægsbærer.
Det fremgår endvidere, at ved et svagt selektionstryk er ændringerne i genfrekvensen altid meget langsomme.
Ved s=1 kan formlen for genfrekvens ændring udvides til at gælde n generationer og man har, når q0 er genfrekvensen i udgangspopulationen:
qn = q0/(1+n*q0)
n = 1/qn - 1/q0
n = 1/0,005 - 1/0,01 = 200 - 100 = 100
Applet til at beregne ændring i genfrekvens for forskellige fitness kombinationer, klik her, se nedenstående graf med eksempel på 1 generations selektion mod den recessive med fitness på 0,5.

DNA test og bortselektion af heterozygote handyr
For recessive sygdomme går der uendelig lang tid med at fjerne et sygdomsgen fra
en population, når man kun kan selektere mod den recessive. I mange populationer
er der relativt meget færre handyr end hundyr, derfor
kan det ofte betale sig kun at DNA teste handyrene. De bidrager med halvdelen
af generne til den nye generation, og man vil derfor kunne nedbringe frekvensen af sygdomsgenet
til halvdelen i hver generation, såfremt der kun avles med hannerne, der er fri for sygdomsgenet.
Man vil samtidig opnå, at der ikke sker nogen udspaltning af de recessive syge individer.
Eksempel: Hos SDM malkekvægracen forekommer den recessivt nedarvede
sygdom, BLAD (Bovin Leucocyte Adhesion Deficiency). I 1992 var man oppe på en genfrekvens
på 15 % i hundyr populationen af BLAD genet. Ved systematisk at
gennemføre 'BLAD DNA test' på de nye tyre er
frekvensen i 1999 langt under 10 %. Og frekvensen af BLAD-genet halveres
for hver ny generation.