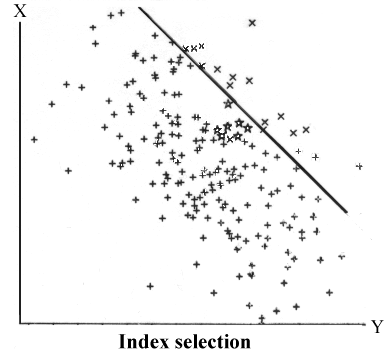
Viser todimensional fordeling med en negativ korrelation på -0,5. Den skrå linie viser selektionsgrænse ved udvalg efter indeks selektion
Effekten af gener i et locus kan påvirke flere egenskaber. Dette medfører, at når der ved selektion sker ændring af en egenskab, vil der, såfremt der er flersidig virkning af de underliggende gener, ske ændring i andre sekundære egenskaber.
Der vil ofte forekomme overraskende resultater. Derfor er det vigtigt, at et selektionsprogram bliver overvåget nøje for at undgå uheldige sekundære ændringer. Er der sammenhæng mellem ændringer i to egenskaber, siger man at de er genetisk korrelerede. Genetisk korrelation kan være positiv eller negativ med talstørrelser fra -1 til +1. Genetisk korrelation har symbolet rA1,A2, hvor A1 og A2 er avlsværdier af de to egenskaber 1 og 2.
Et eksempel fra kødkvægavl. Der selekteres for store kalve ved fødsel - såfremt køernes udvoksede vægt ikke ændres samtidig, vil det uvægerligt resultere i kælvningsbesvær, idet forholdet mellem kalv og koen, der skal føde, ændres i ugunstig retning.
| Figur 8.6.
Viser todimensional fordeling med en negativ korrelation på -0,5. Den skrå linie viser selektionsgrænse ved udvalg efter indeks selektion |
|
I figur 8.6 er vist en todimensional fordeling med negativ genetisk korrelation på 0,50 mellem egenskaberne X og Y. Selektionsgrænse (den skrå linie) er lagt, så der er lagt lige meget vægt på de to egenskaber. Som det ses, er det svært at forbedre begge egenskaber ret meget når der er negativ genetisk korrelation. Omvendt går det meget lettere, såfremt egenskaberne har positiv genetisk korrelation i forhold til den ønskede selektionsretning.
Der tales om tre forskellige udvalgskriterier
Tandem selektion foregår med en selektion af egenskab ad gangen
Uafhængig selektion, her sættes selektionsgrænserne
uafhængigt for hver egenskab
Indeks selektion, her vægtes egenskaberne i forhold til deres økonomiske betydning
Det er svært at bestemme den genetiske korrelation, idet den kan variere meget fra population til population, og den varierer meget mere over tid end heritabiliteten. Der skal dog gives en formel til beregning af realiseret genetisk korrelation. Når der selekteres for egenskaben x og der samtidig sker ændring i egenskaben y, kan den genetiske korrelation bestemmes som:
rAx,Ay = (delta Gy/sigmaAy)/(delta Gx/sigmaAx)
Den genetiske korrelation bestemmes altså som forholdet mellem de standardiserede selektions resultater, når der kun selekteres for egenskaben x, og egenskaben y følger passivt med.
Der er en tendens til at der ved selektion gennem mange generationer vil opstå en uheldig genetisk korrelation mellem den egenskab, man selekterer på og 'fitness'. Dette giver sig udslag i, at der hele tiden skal være et selektionstryk på egenskaben for at holde den på det niveau, hvortil man er nået. Opgives selektionen, vil populationen gå tilbage mod udgangspunktet hvor der er optimal fitness.
| Figur 8.7.
Viser at ved store ændringer af en egenskab er det ofte også nødvendig at ændre miljøforholdene for at opnå maksimal effekt (læses harmoniske dyr) ved udnyttelse af interaktionen. |
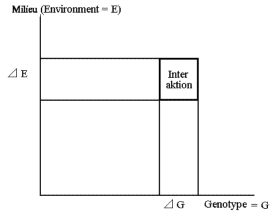 |
Ved store ændringer både i milieu-forhold eller genetisk baggrund, bliver interaktionen mellem genetik og milieu en nødvendig faktor at forholde sig til. Interaktionen afspejler den negative korrelation til fitness.
Selektionsforsøg til belysning af genetisk korrelation. I figur 8.8 er vist tre parallelle selektionsforsøg med
slagtekyllinger gennemført af Australieren Pym, der har offentliggjort resultaterne i 1982. Forsøget gik over 10 generationer, og måleperioden var fra 5 til 9 ugers alderen. der er selekteret for henholdsvis Weight gain (W), Food consumption
(F) og Food conversion rate (E) i de tre forsøg, og kurverne for de tre egenskaber har betegnelserne W, F og E.
Samtidig med at man har målt det direkte selektionsrespons, har man også målt, hvordan det er gået med de to andre
egenskaber. At de ikke-selekterede egenskaber også ændrer sig er udtryk for genetisk korrelation. Det kan endvidere ses, at
selektionen resulterer i vidt forskellige kyllingetyper afhængigt af, hvilken egenskab der selekteres for.
| Figur 8.8.
Selektionsforsøg med slagtekyllinger til belysning af genetisk korrelation gennemført af Australieren Pym. Forsøget gik over 10 generationer, og måleperioden var fra 5 til 9 ugers alderen. der er selekteret for henholdsvis Weight gain (W, øverste graf)), Food consumption (F, mellemste graf) og Food conversion rate (E, nederste graf). De korrelerede resultater er vist i de andre grafer, så F i øverste graf refererer til korreleret respons for W ved selektion for F, o.s.v.. |
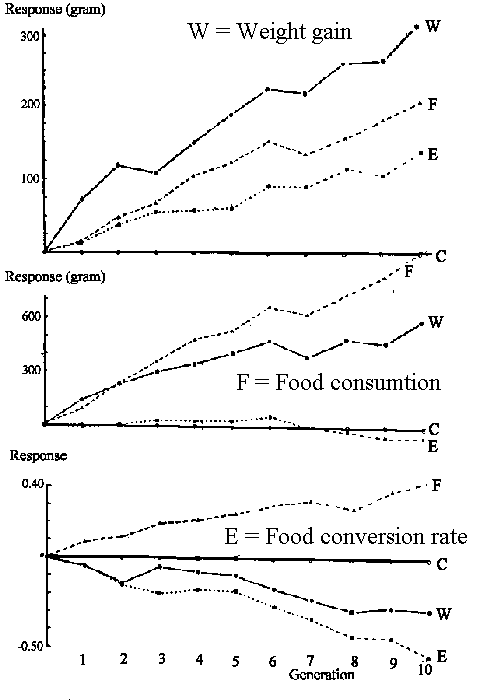 |
Dyr der selekteres for tilvækst (øverste graf W) får også et større fødeindtag
(mellemste graf W) og en bedre effektivitet (nederste graf W) lavere foderforbrug
per kg tilvækst.
Dyr der selekteres for foderforbrug (mellemste graf F) får også en større tilvækst
(øverste graf F) og en dårligere effektivitet (nederste graf F) højere foderforbrug
per kg tilvækst.
Dyr der selekteres for effektivitet (nederste graf E) får også en større tilvækst
(øverste graf E) og et uændret foder indtag (mellemste graf E).
Video klip af stærkt selekterede kyllinger for dgl. tilvækst
med korrelerede benproblemer,
(kræver installation af Real Audio)
Video klippene stammer fra en udsendelse fra DR 'Journalen' og er optaget på
Danmarks Jordbrugsforskning, Foulum, hvor prof. Poul Sørensen har stået for
den faglige information.
Video klip
af kyllinger, der er selekteret for stor dgl. tilvækst, og en tilsvarende æglægger. Ved
5 uger vejer den hvide slagtekylling ca. 2 kg, der er ca 4 gange vægten af den brune
æglægger kylling.
Video klip
af kyllinger der er selekteret for stor dgl. tilvækst, der som korreleret effekt får
benproblemer. Den først viste kylling har så svære benproblemer, at den ikke kan
klare at få føde og vand, den bør aflives. Kylling nummer to går dårlig, og den
har gang som en sømand der ved at skræve får balance i kroppen.
Auto selektion for egenskaben kuldstørrelse - balance med fitness.
| Figur 8.9.
Viser kuldstørrelse som egenskab for søerne og som egenskab hos de fødte grise, der er som vist 1 gris til forskel mellem de to middelværdier, der svare til en selektionsdifferens på 1 i hver generation. |
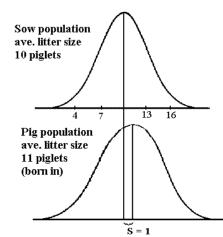 |
Der er i hver generation en auto selektion for højere kuldstørrelse, da der bliver født flest grise i de store kuld. Det kan vises, at der er en selektions- differens på en gris for højere kuldstørrelse i hver generation, se figur 8.9. Egenskaben kuldstørrelse er normal fordelt når man ser på egenskaben fra søernes side, men egenskaben født i kuldstørrelse er højreskæv, da der som før nævnt bliver født flest grise i de store kuld. Denne selektion må nødvendigvis balanceres af at de grise der er født i store kuld har lavere fitness end dem der er født i små kuld.
Den maternelle påvirkning for en grises videre skæbne er positiv for grise født i små kuld og negativ for grise født i store kuld. Ved udtagning af sopolte bør man derfor undgår individer født i store kuld. Den negative maternelle effekt gør sig især gældende i dyrearter, hvor kønsmodenhed indtræder før dyrene er udvoksede.
Tilbageslag efter ophørt selektion for egenskaber hvori fitness indgår.
Såfremt man ved selektion har ændret en egenskab meget vil den ofte gå tilbage mod
udgangspunktet når selektionen ophører. F.eks. tamsvin har en kuldstørrelse omkring 10,
hvorimod deres stamfædre, vildsvinet, kun har en kuldstørrelse på det halve. Man kan
derfor forvente, at såfremt man lader tamsvinet gå tilbage til naturtilstanden vil
kuldstørrelsen efterhånden tilpasse sig til de forhåndenværende ressourcer. De samme
synspunkter vil også være gældende for højtydende malkekøer såvel som for æglæggere eller
slagtefjerkræ.
De højt specialiserede husdyrracer vil kun kunne opretholde den store produktion,
i det omfang der bliver sørget for dem hvad angår indhusning og fodertildeling.