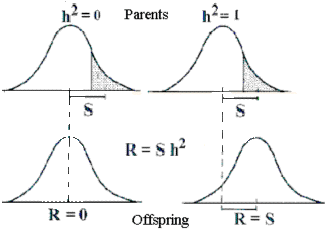
Selektionsresultatet (R) er lig heritabiliteten gange selektionsdifferensen (S) Ændringen ses hos afkommet
Når der tales om additiv varians refererer det til varians på avlsværdierne (VA). (I senere lektioner anvendes ofte symbolet sigma2 som synonym for varians.) I det foregående er der vist, hvorledes avlsværdi kan beregnes for de enkelte genotyper i et locus. Når værdierne er kendte, kan der også beregnes en varians på avlsværdierne i dette locus. Såfremt der er uafhængighed mellem de enkelte loci, der har additiv effekt på en egenskab, bliver den samlede additive varians summen af varianser fra de enkelte loci. Den samlede additive varians kan ikke umiddelbart beregnes for individer i en population, idet avlsværdien for de enkelte individer ikke er kendt.
Den phenotypiske varians (VP) derimod beregnes på grundlag af målinger på de enkelte individer.
Heritabiliteten (h2) eller arvelighedsgrad for en egenskab er defineret som VA/VP. Heritabiliteten varierer derfor mellem 0 og 1, idet den additive varians er en del af den phenotypiske varians.
Beregning af heritabilitet kan ske på grundlag af lighed mellem beslægtede individer, målt som korrelation eller som regression, f.eks. regression af afkom på en af forældrene. Og der gælder følgende relation. Heritabiliteten kan beregnes som forholdet mellem den beregnede korrelation og slægtskabskoefficienten. Dvs. den maksimale korrelation mellem forældre og afkom er på en halv. Såfremt den er højere, må der være andre end genetiske årsager til at forældre og børn ligner hinanden. Den teoretiske baggrund for den forventede additive kovarians mellem beslægtede individer kan udledes efter lignende ræsonnementer som for den additive varians, idet man først beregner kovariansen i et enkelt locus og derefter summerer over alle loci.
Den bedste metode til beregning af heritabilitet er korrelation mellem halvsøskende, da de normalt kun har gener til fælles. Helsøskende derimod har ofte et maternelt milieu fælles, hvilket i så fald fører til overestimering af heritabiliteten. I praksis bruges sjældent forældre-afkom beregning af korrelation, da man så nødvendigvis skal skaffe data, der stammer fra to generationer. Der er endvidere problemer med at forældre og afkom ikke lever under de samme milieuvilkår, eller evt. at forældre og afkom deler fælles milieu.
En estimering af variansandelen, der skyldes fælles milieu, kan ske ved at sammenligne korrelation beregnet på grundlag af halv- og helsøskende. Forventningen er, at korrelation mellem helsøskende skal være dobbelt så stor som korrelation mellem halvsøskende. Det faktum, at den er større end to gange halvsøskende korrelationen, kan tilskrives fælles milieu for helsøskende. Denne komponent får betegnelsen (c2), hvor c står for 'common environment'. Den relative phenotypiske varians består af en genetisk del (h2) fælles milieu variansen (c2). Den resterende del af variansen skyldes tilfældige milieupåvirkninger, og den andel får betegnelsen (e2). For de fleste egenskaber udgør milieuvariansen den største del af den phenotypiske varians.
Tabel over de definerede genetiske parametre og deres symboler Additiv varians VA Phenotypiske varians VP Heritabiliteten h2 Fælles milieu c2 Tilfældig milieu e2 Halvsøskende korrelation (1/4)h2
| Figur 6.3.
Selektionsresultatet (R) er lig heritabiliteten gange selektionsdifferensen (S) Ændringen ses hos afkommet |
|
|
Historisk set har heritabiliteten en fortolkning mere. Såfremt der sker selektion, så de udvalgte avlsdyr (skraveret område i figur 6.3) afviger fra populationsgennemsnittet, vil gennemsnittet i næste generation ændres. Ændringen er proportional med selektionen og heritabiliteten og man har følgende formel:
R = h2*S
Selektions-resultatet eller -respons (R) er lig heritabiliteten gange selektionsdifferensen (S). Heritabiliteten er nul, såfremt egenskaben forbliver konstant ved selektion, og den er 1, såfremt ændringen er lige så stor som selektionsdifferensen, se figuren til højre og lektion 1 med ændring af fedtprocenten i mælken hos Dansk Jersey kvæg på grundlag af selektion.